
jueves, 5 de abril de 2012
Digestión de proteínas
La digestión de proteínas comienza en el estómago. La
entrada de proteínas al estómago estimula la secreción de gastrina,
la cual a su vez estimula la formación de HCl; esta
acidez actúa como un antiséptico y mata a la mayoría de los entes patógenos que
ingresan al tracto intestinal.
En el
estómago, la pepsina, de una sola cadena, es secretada en forma de su
zimógeno (enzimas inactivas) , el pepsinógeno por las células de la mucosa gástrica. El pepsinógeno
se convierte en pepsina por el corte (catalizado por
la misma enzima) de 42 residuos del extremo amino-terminal, proceso que es
favorecido por el pH ácido del jugo gástrico. La pepsina no es muy específica,
hidroliza los enlaces en los que intervienen aminoácidos aromáticos, aunque
también lo hace donde hay Met y Leu.
A medida que
los contenidos ácidos del estómago pasan al intestino delgado, se dispara la
síntesis de la hormona secretina a la sangre. Esta enzima estimula al páncreas
para secretar bicarbonato en el intestino delgado para neutralizar el pH
alrededor de 7.0. La entrada de los aminoácidos en la parte superior del
intestino (duodeno) se libera la hormona colecistocinina,
que estimula la liberación de muchas enzimas pancreáticas cuya actividad
catalítica se realiza entre 7 y 8 unidades de pH. El jugo pancreático secretado
al intestino delgado aporta los zimógenos de
tripsina, quimotripsina, tripsinógeno,
carboxipeptidasas A y B y elastasa.
Por ejemplo,
el quimotripsinógeno da origen a la quimotripsina por
separación de 2 dipéptidos. Su conversión
a alfa-quimotripsina se debe a la hidrólisis enzimática de 4 enlaces peptídicos
por acción de la tripsina y quimotripsina
consecutivamente:
La
pancreatitis, condición dolorosa y a menudo fatal, se caracteriza por la
activación prematura de proteasas secretadas por el
páncreas.
El tripsinógeno , da origen a la tripsina por
separación de un hexapéptido del amino-terminal por
acción de la enterocinasa. La tripsina hidroliza
enlaces en los que intervienen Arg y Lys.

Enzima de escape
La enzima que se detecta en el infarto agudo al miocardio es la CKmb
El ácido glutamico y el oxaloacetato por acción de la TGO (TRANSAMINASA GLUTAMICO OXALOACETICA) intercambiaran sus grupos funcionales para formar alfa cetoglutarato y aspartato.
Esto también ocurre a la inversa, es decir , que alfa cetoglutarato y aspartato intercambiaran sus grupos funcionales por acción de la ASAT ( TRANSAMINASA ASPARTICO AMINOTRANSFERASA) y formaran el ácido glutamico y el oxaloacetato
El ácido glutamico y el piruvato por acción de la TGp (TRANSAMINASA GLUTAMICO PIRUVICA) intercambiaran sus grupos funcionales para formar alfa cetoglutarato y alanina. Esto también ocurre a la inversa puesto que alfa cetoglutarato y alanina intercambian sus grupos funcionales por la acción de ALAT (ALANINA AMINOTRANSFERASA) forman ácido glutamico y el piruvato
Moléculas derivadas de los aminoácidos
1) Las porfirinas que se forman a partir de la glicina son la base estructural del grupo hemo de la hemoglobina y la mioglobina
2) A partir de glicina y Arginina se sintetiza fosfocreatina la cual es una forma de almacenamiento de energía en los músculos
3) La histamina se origina de la histidina, esta involucrada en la inflamación, desencadena vasodilatación, vasoconstricción y movilización de células.
4) La tirosina es el precursor de la melanina, las catecolaminas y las hormonas de la tiroides. La melanina es un pigmento del la piel y el pelo, que protege de los rayos ultravioleta. Las catecolaminas son un grupo de compuestos como la Dopamina, neurotrasmisor cerebral relacionado con las funciones motrices; la adrenalina,
hormona secretada en situaciones de alerta que aumenta la glucemia, el ritmo cardiaco y la presión arterial; y la noradrenalina que desempeña funciones parecidas a la adrenalina. Las hormonas tiroxina y triyodotironina estimulan el metabolismo de los carbohidratos, lípidos y aminoácidos.
hormona secretada en situaciones de alerta que aumenta la glucemia, el ritmo cardiaco y la presión arterial; y la noradrenalina que desempeña funciones parecidas a la adrenalina. Las hormonas tiroxina y triyodotironina estimulan el metabolismo de los carbohidratos, lípidos y aminoácidos.
5) La serotonina y la melatonina se forman a partir de triptófano. La serotonina es un neutrotrasmisor, induce el sueño, controla el apetito, inhibe la secreción gástrica, aumenta el peristaltismo, estimula la secreción de hormonas de la hipófisis, produce vasoconstricción, disminuye la contracción del corazón, aumenta la agregación plaquetaria y es broncoconstrictor. La melatonina al igual que la serotonina modula el sueño, además controla los ciclos reproductivos de acuerdo al fotoperiódo, recientemente se ha relacionado con el envejecimiento.
Transaminaciones
Para las transaminaciones se requiere de la vitamina B6 o también conocida como priridoxina y en el metabolismo se encuentra como prifofosfato de piridoxina
Los dos aminoácidos que son cetógenicos exlusivamente son leucina y lisina.

Los dos aminoácidos que son cetógenicos exlusivamente son leucina y lisina.

Metabolismo del ácido araquidónico
Fosfatidilinositol que es un glicerofosfolípido por accion de la fosfolipasa A2 que provoca una ruptura en el carbono 2 y este libera Ac Araquidónico.
El ácido araquidónico es una ácido graso insaturado.
Por acción de la E. Ciclooxigenasa para formar eicosanoides conocidos como: leucotrienos, tromboxanos y prostaglandinas.
Dichos eicosanoides son los causantes de la fiebre, coagulación y algesia respectivamente.
Es por ello que la aspirina, naproxeno, ketorolaco, diclofenaco son analgesicos pues inhiben a la ciclooxigenasa

El ácido araquidónico es una ácido graso insaturado.
Por acción de la E. Ciclooxigenasa para formar eicosanoides conocidos como: leucotrienos, tromboxanos y prostaglandinas.
Dichos eicosanoides son los causantes de la fiebre, coagulación y algesia respectivamente.
Es por ello que la aspirina, naproxeno, ketorolaco, diclofenaco son analgesicos pues inhiben a la ciclooxigenasa

Fosfolípidos
Clasificación de los Fosfolípidos


1) Los glicerofosfolípidos cuentan con un ac, graso saturado y un ac. graso insaturado más un grupo funcional y se dividen en:
a) fosfatidiletanolamida: principal componente de las memranas celulares y se dividen en Plasmalógeno que forma el 70 % de las membranas celulares y el aquilacifosfolípido que forma parte d elas membranas celulares de los eritrocitos
b) Fosfatidilcolina: forma parte del factor surfactante producido por los neumocitos tipoII
c)Fosfatidilserina: su precursor es la fosfatidiletanolamida y funciona como 2° mensajero y neurotransmisor
d) Fosfatidil inositol: Interviene en el metabolismo del ac Araquidónico
e)Fosfatidilglicerol: Cardiolipina (forma parte de las membranas celulares del corazón)
2.- Los esfingolípidos se dividen en tres familias cerebrósidos, globósidos y gangliósidos
Degradación de bases pirimidinicas
1) Entra la CMP y la UMP por accion de una E. nucleotidasa se forma citidina y uridina respectivamente
2) la CITIDINA por acción de la E. citidina desaminasa se convierte en Uridina.
3) La uridina por accion de la E. uridina fosforilasa se convierte en Uracilo
4) Uraciolo por acción de la E. Dihidrouracilo deshidrogenasa se convierte en dihidrouracilo
5)Dihidrouracilo por accion de la E. hidropirimina hidratasa se forma ureidopropionato
6) Ureidopropionato por acción de la E. Ureidopropionasa se forma B alanina
7) B alanina por acción de la E. aminotransferasa se forma semilaldehído malónico
8) Semialdheído malónico de origen a malonil co A.
Malonil coA puede entrar a la via de la B- Reducción
2) la CITIDINA por acción de la E. citidina desaminasa se convierte en Uridina.
3) La uridina por accion de la E. uridina fosforilasa se convierte en Uracilo
4) Uraciolo por acción de la E. Dihidrouracilo deshidrogenasa se convierte en dihidrouracilo
5)Dihidrouracilo por accion de la E. hidropirimina hidratasa se forma ureidopropionato
6) Ureidopropionato por acción de la E. Ureidopropionasa se forma B alanina
7) B alanina por acción de la E. aminotransferasa se forma semilaldehído malónico
8) Semialdheído malónico de origen a malonil co A.
Malonil coA puede entrar a la via de la B- Reducción
Degradación de bases Púricas
Los nucleótidos AMP y GMP, o sus análogos, pierden su grupo fosfato por acción de la enzima nucleotidasa con lo que se producen los nucleósidos adenosina y guanosina.
La adenosina es desaminada por la adenosina desaminasa y se convierte en inosina. A continuación guanosina e inosina pierden su pentosa gracias a la nucleósido de purina fosforilasa, en consecuencia quedan libres las purinas guanina e hipoxantina respectivamente.
Ambas se convierten en xantina por acción de las enzimas guanina desaminasa y xatina oxidasa, esta última también es se encarga posteriormente de transformar a la xantina en ácido úrico.
La xantina oxidasa es la reguladora de la vía y puede ser inhibida por alopurinol y colchicina. Estas se utilizan en el trtamiento de la gota una enfermedad producida por el aumento de ácido úrico y produce inflamación en las articulaciones distales

La adenosina es desaminada por la adenosina desaminasa y se convierte en inosina. A continuación guanosina e inosina pierden su pentosa gracias a la nucleósido de purina fosforilasa, en consecuencia quedan libres las purinas guanina e hipoxantina respectivamente.
Ambas se convierten en xantina por acción de las enzimas guanina desaminasa y xatina oxidasa, esta última también es se encarga posteriormente de transformar a la xantina en ácido úrico.
La xantina oxidasa es la reguladora de la vía y puede ser inhibida por alopurinol y colchicina. Estas se utilizan en el trtamiento de la gota una enfermedad producida por el aumento de ácido úrico y produce inflamación en las articulaciones distales

Síntesis de bases pirímidinicas
La enzima reguladora de la vía es la carbamoil fosfato sintetasa II
Las bases pirimidinicas son:



En este caso el anillo de pirimidina se sintetiza primero y por separado, posteriormente se une al PRPP para dar lugar al monofosfato de orotidina (OMP), a partir del cual se forman los otros nucleótidos en forma secuencial.
Los sustratos para la construcción de estos nucleótidos son el carbamoil fosfato, aspartato y PRPP, el carbamoil fosfato se forma en los animales por la condensación en el citosol de CO2 y el ión amonio gracias a la carbamoil sintetasa II.
Éste y el aspartato dan lugar al carbamoil aspartato que se transforma en dihidrooroato y finalmente en oroato, que es el primer metabolito que contiene el anillo de pirimidina.
Luego el oroato se une al PRPP produciéndose monofosfato de orotidina, el cual se descarboxila y genera monofosfato de uridina (UMP).
De manera semejante a lo que ocurre con los nucleótidos de purina, el trifosfato de uridina (UTP) se forma por fosforilación del UMP, aunque en este caso se realiza en un sólo paso con el gasto de 2 ATP. El trifosfato de citidina (CTP) se sintetiza a partir del UTP, el nitrógeno extra que contienen el anillo de citosina proviene de la
glutamina, este paso requiere además de la energía proveniente de un ATP.
Las bases pirimidinicas son:



En este caso el anillo de pirimidina se sintetiza primero y por separado, posteriormente se une al PRPP para dar lugar al monofosfato de orotidina (OMP), a partir del cual se forman los otros nucleótidos en forma secuencial.
Los sustratos para la construcción de estos nucleótidos son el carbamoil fosfato, aspartato y PRPP, el carbamoil fosfato se forma en los animales por la condensación en el citosol de CO2 y el ión amonio gracias a la carbamoil sintetasa II.
Éste y el aspartato dan lugar al carbamoil aspartato que se transforma en dihidrooroato y finalmente en oroato, que es el primer metabolito que contiene el anillo de pirimidina.
Luego el oroato se une al PRPP produciéndose monofosfato de orotidina, el cual se descarboxila y genera monofosfato de uridina (UMP).
De manera semejante a lo que ocurre con los nucleótidos de purina, el trifosfato de uridina (UTP) se forma por fosforilación del UMP, aunque en este caso se realiza en un sólo paso con el gasto de 2 ATP. El trifosfato de citidina (CTP) se sintetiza a partir del UTP, el nitrógeno extra que contienen el anillo de citosina proviene de la
glutamina, este paso requiere además de la energía proveniente de un ATP.
Sintesis de bases Púricas
El alimentador de esta vía es la Ribosa-5 fosfato producida en la Vía de las pentosas fosfato.
Las bases púrcas son: Adenina y Guanina


La nucleósido fosforilasa rompe el enlace glucosídico entre la ribosa y la base nitrogenada, mediante la adición de fosfato inorgánico dando lugar a ribosa 1-fosfato más la base correspondiente. La nucleósido fosforilasa también cataliza la reacción en forma inversa, es decir, el primer paso en la síntesis de salvamento de nucleótidos a partir de bases nitrogenadas libres. Cuando esto ocurre, el nucleósido producido
puede fosforilarse por el ATP mediante una nucleósido quinasa. Estas enzimas no son universales, por ejemplo, los animales no contienen guanosina quinasa. La formación de los nucleótidos a partir de las bases nitrogenadas liberadas y del 5- fosfo--D-ribosil-1-pirofosfato (PRPP), sintetizado en la célula, es otra alternativa en la ruta de salvamento, esta reacción es catalizada por la fosforribosiltransferasa.
El PRPP además es un intermediario muy importante en la síntesis de novo de nucleótidos de purina y pirimidina. En la síntesis de novo primero se forman los ribonucleótidos y posteriormente los desoxirribonucleótidos. El primer ribonucleótido de purina totalmente formado es la 5´monofosfato de inosina (IMP) o ácido inosínico. Su doble anillo se va formando poco a poco sobre el PRPP, de sus cuatro átomos de nitrógeno dos provienen de la glutamina, uno de la glicina y otro del aspartato. Se requieren de cuatro ATP y de tetrahidrofolato. Este último es una coenzima proveniente de la vitamina folacina.
El IMP origina monofosfato de guanosina (GMP) y monofosfato de adenosina (AMP) en vías separadas. En el primer caso, el anillo de purina de la IMP se hidroxila en una reacción dependiente de NAD produciendo monofosfato de xantosina, que después se convierte en GMP por una amidotrasferasa dependiente de glutamina y ATP. La síntesis de AMP involucra la transferencia de nitrógeno desde el aspartato. En primer lugar se genera adenilosuccinato como intermediario y a continuación por eliminación de fumarato se libera AMP. La energía que impulsa esta reacción no proviene del ATP, sino del GTP.
El GMP y el AMP se convierten sus correspondientes trifosfatos a través de dos reacciones de fosforilación sucesivas. La conversión a difosfatos es catalizada por quinasas dependientes de ATP. La fosforilación de ADP en ATP se produce principalmente en la fosforilación oxidativa. Luego el ATP es el donador de fosfato para la conversión del GDP y otros dinucleótidos, a sus formas trifosfatadas.
Ciclo de la urea
En animales amoniotélicos,
por ejemplo, peces óseos, el amoníaco se libera rápidamente de la
sangre en las branquias, gracias al gran volumen de agua que pasa a
través de éstas. Las bacterias y protozoos simplemente liberan el
amoníaco al medio en que el agua es abundante, donde se disuelve este
compuesto
Mientras que los animales uricotélicos,
las aves y reptiles, la disponibilidad de agua es limitada. Puesto que
la excreción de urea por la orina necesita un gran volumen de agua, esta
circunstancia haría imposible el vuelo de las aves y provocaría una
deshidratación de los reptiles que habitan hábitats áridos. Para evitar
esto, el amoniaco se convierte en ácido úrico, compuesto insoluble que
se excreta en forma de masa semisólida de cristales de ácido úrico en
las heces.
En la especie humana, ureotélica,
el ión amonio es un compuesto muy tóxico que se convierte en el hígado y
el riñón en urea, en el llamado ciclo de la urea. Ésta pasa al torrente
sanguíneo y es eliminada por el riñón en la orina.

La acumulación de amonio tiene consecuencias tóxicas. Por lo tanto se debe eliminar con la misma rapidez con la con la que se genera. Los animales acuáticos eliminan directamente amonio gracias a que pueden captar y expulsar cantidades ilimitadas de agua. En cambio los animales terrestres necesitan
trasformarlo en un compuesto que pueda excretarse sin que ello implique una pérdida importante de agua. Las aves y los reptiles producen ácido úrico y los mamíferos urea. La urea se forma a lo largo de una secuencia de cinco reacciones en el hígado, de la cuales cuatro forman un ciclo:
1) En la mitocondria, la enzima mitocondrial cabamoil fosfato sintetasa I, que técnicamente no forma parte del ciclo de la urea, cataliza la reacción limitante. Condensa amonio y bicarbonato para formar carbamoil fosfato, quien proporcionauno de los dos átomos de nitrógeno de la urea. Esta reacción es irreversible y
requiere de 2 ATP. En los eucariotas la carbamoil fosfato sintetasa II es citosólica, usa glutamina como donador de nitrógeno y esta involucrada en la síntesis de pirimidinas.
2) El grupo carbamoil del carbamoil fosfato es transferido a la ornitina formando citrulina. Esto ocurre dentro de la mitocondria gracias a la ornitinatranscarbamoilasa. La citrulina debe salir de la mitocondria para que pueda continuar el ciclo de reacciones.
3) La citrulina, en el citosol, se condensa con el aspartato produciendo argininsuccinato, el aspartato proporciona el segundo átomo de nitrógeno de la urea. La argininsuccinato sintetasa, responsable de la reacción, requiere de dos enlaces de alta energía del ATP.
4) El argininsuccinato se convierte en arginina al liberar fumarato, con la participación de la argininosuccinasa. La arginina es el precursor inmediato de la urea. En el ciclo de Krebs el fumarato se transforma en oxalacetato, el cual por transaminación se convierte nuevamente en aspartato.
5) Por último la arginasa hidroliza a la arginina con lo que se restaura la ornitina y se libera la urea. La urea es excretada a través de la orina y la ornitina es trasladada a la mitocondria, para que nuevamente reaccione con el carbamoil fosfato y el ciclo continúe.
domingo, 1 de abril de 2012
Colesterogenesis

1. Acetil CoA pasa a Acetoacetil CoA por una enzima tiolasa
2. Acetoacetil CoA pasa a B-hidroxi-B-metilglutaril CoA por la hidroximetilglutaril CoA sintasa
3. B-hidroxi-B-metilglutaril CoA pasa Mevalonato por la hidroximetilglutaril CoA reductasa que es la enzima reguladora de la colesterogenesis
Estos pasos ocurren en el citoplasma de la célula hepática

4. El mevalonato pasa a 5 fosfomevalonato por la mevalonato fosfotransferasa o mevalonato cinasa
5. 5fosfomevalonato pasa a 5 pirofosfomevaloato por la 5 fosfomevalonato cinasa
6. 5 pirofosfomevalonato pasa a 3 fosfo-5 pirofosfomevalonato por la 5 pirofosfomevalonato descarboxilasa
7. 3 fosfo-5 pirofosfomevalonato se descarboxila y da los isoprenoides activados isopentenil pirofosfato (1° unidad isoprenide) y dimetalil pirofosfato (2° unidad isoprenoide)

{kind=link}
8. La unión de las dos unidades isoprenoides forma el terpeno geranil pirofosfato (10C)
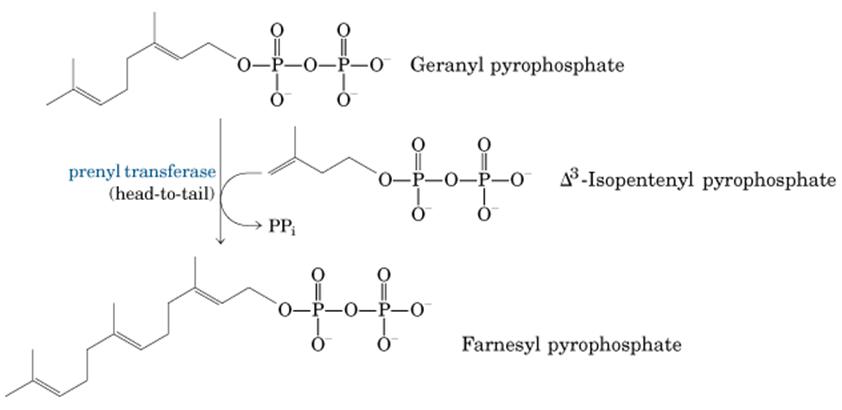
9. La prenil transferasa añade una unidad isoprenide (5C) al geranil pirofosfato y da al terpeno farneisl pirofosfato (15C)
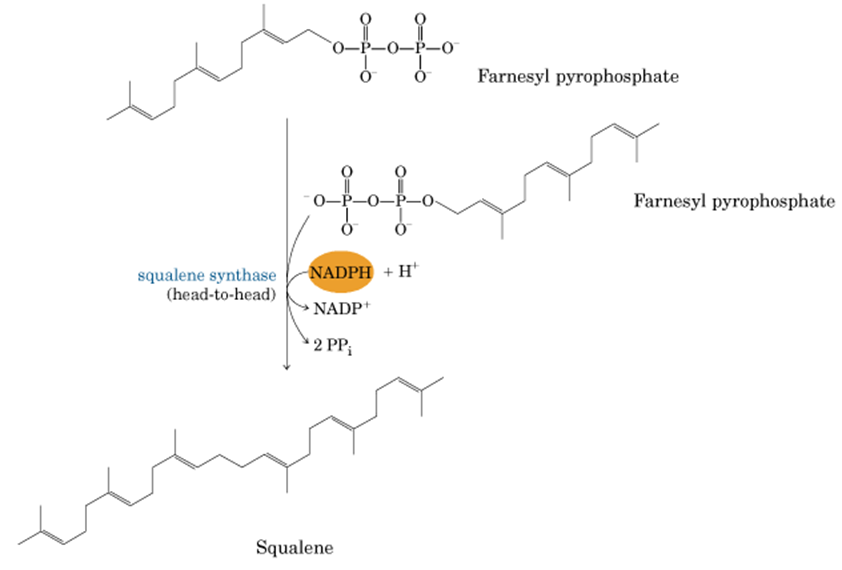
10. La unión de dos moléculas de farnseil pirofosfato por la escualeno sintasa da al escualeno

11. El escualeno pasa por una serie de reacciones donde llega a ser una molécula de colesterol
Síntesis de cuerpos cetonicos

Esta vía se realiza en la mitocondria
1. Acetil CoA alimenta la vía y se convierte en aceto acetil CoA por una enzima tiolasa
2. Acetoacetil CoA pasa a B-hidroxi-B-metilglutaril- CoA por la enzima hidroximetilglutaril CoA sintetasa (HMGCoA)
3. B-hidroxi-B-metilglutaril CoA pasa a acetoacetato (!° cuerpo cetonico) por la HMG CoA liasa
4. Acetoacetato pasa a acetona (2° cuerpo cetonico) por acetoacetato descarboxilasa
5. Acetoacetato pasa a D-B hidroxibutirato por D-B hidroxibutirto deshidrogenasa
Beta oxidación
Es la vía por la cual se degradan ácidos grasos



1. Paso de acil CoA (palmitoil CoA) a trans delta enoil CoA por la enzima acil CoA deshidrogenasa donce el cofactor FAD acepta 2H
2. Trans delta enoil CoA pasa LB hidroxiacil CoA por la trans delta enoil CoA hidrolasa
3. LB hidroxiacil CoA pasa a B cetoacil CoA por la LB hidroxiacil CoA deshidrogenasa
4. B cetoaacil CoA forma Acil CoA (miristiril CoA) y Acetil CoA por la acil CoA acetil transferasa

Esteroides
Los esteroides son lípidos de la más alta importancia en la fisiología humana y su estructura química deriva del núcleo del ciclopentano perhidrofenantreno. Un grupo formado por los tres anillos del fenantreno pero con sus dobles enlaces saturados, unido al ciclopentano. Este grupo químico que es característico de todos los esteroides se modifica son varios sustituyentes alcohol o cetona en diversas posiciones de los anillos y también por una cadena de carbonos unida al carbono 17 del ciclopentano (C-17).
El colesterol, molécula de 27 carbonos -cuyo nombre significa “alcohol sólido de la bilis” pues se encuentra con alguna frecuencia formando cálculos biliares radiolúcidos de apariencia opalina- es el compuesto original que da lugar a la formación de los diferentes esteroides, los cuales en número de varias decenas intervienen en las funciones del organismo humano, la mayoría de ellos como hormonas; pero también en función de vitaminas y de agentes tensoactivos.
Además de ser el precursor de todos los esteroides, el colesterol mismo tiene funciones importantes en el organismo, entre ellas la de formar parte de las membranas y la de participar en la cubierta monocapa de las lipoproteínas. Su acarreo en la circulación por las lipoproteínas LDL es motivo del interés clínico pues se le asigna el papel principal en la génesis de la aterosclerosis, alteración patológica que -en mayor o menor grado- ocurre de manera universal en todos los seres humanos y que consiste en la formación de “placas de ateroma” constituidas principalmente por el depósito de colesterol en la íntima de las medianas y las grandes arterias.
Desde el punto de vista de su estructura química, los esteroides se pueden dividir en cuatro categorías según el número de carbonos insertos en la cadena lateral del C-17:
a)- 8 carbonos: Esteroles, por ejemplo el colesterol y la vitamina D, además de otros esteroles vegetales como el sitosterol y el estigmasterol.
b)- 5 carbonos: Ácidos biliares y sus sales, por ejemplo los ácidos cólico, glicocólico, taurocólico, desoxicólico y litocólico.
c)- 2 carbonos: Progesterona y esteroides de las suprarrenales, glucocorticoides como la cortisona y el cortisol y mineralocorticoides como la desoxicorticosterona (DOCA) y la aldosterona.
d)- 0 carbonos: Hormonas sexuales masculinas y femeninas, testosterona y estradiol.

Clasificación de lípidos
LÍPIDOS SIMPLES:
TRIACILGLICEROLES
También llamados triglicéridos o grasas neutras, son los lípidos más abundantes en los organismos vivos y están formados por el alcohol glicerol esterificado con tres ácidos grasos. Las moléculas de triacilgliceroles en las grasas naturales son muy variadas pues cada uno de los tres ácidos grasos puede ser alguno de los cerca de 10 ácidos grasos más frecuentes, lo cual hace posible las características observadas en la grasa de las distintas especies, por ejemplo: el sebo, la manteca, la mantequilla y los aceites.
Las principales funciones de los triacilgriceroles es la de constituir la reserva más grande de energía en el organismo humano y la única que permite la sobrevida durante el ayuno prolongado y la función nutricional pues las grasas figuran en la dieta diaria aportanto alrededor del 30% de las kilocalorias necesarias para el mantenimiento del organismo; cada gramo de grasa aporta 9 Kcal.
Las grasas corporales funcionan también como amortiguador mecánico para proteger a los tejidos, por ejemplo: la grasa que rodea a los riñones, el corazón y el intestino. La grasa subcutánea protege también al cuerpo de los agentes mecánicos externos y además funciona como un aislante térmico que protege a los organismos de las bajas temperaturas.

ÁCIDOS GRASOS
Son ácidos monocarboxílicos de cadena lineal R-COOH, donde R es una cadena alquilo formáda sólo por átomos de carbono e hidrógeno. Como parte de los triglicéridos existen más de 20 ácidos grasos diferentes. La longitud de la cadena de carbonos varía entre 4 y 24 aunque los más comunes contienen 16 o 18 átomos de carbono. Además de la longitud, la cadena carbonos puede ser saturada o insaturada, es decir, que tiene generalmente de uno a cuatro dobles enlaces carbono-carbono. La insaturación de los ácidos grasos repercute en las propiedades físicas de la grasa pues los ácidos grasos insaturados tienen puntos de fusión más bajos que los saturados correspondientes. Las grasas que tienen en su mayoría ácidos grasos saturados son sólidas o semisólidas a temperatura ambiente: sebo de res o de cordero, manteca de cerdo, la mantequilla o la margarina; en cambio los aceites que son líquidos a temperatura ambiente están formados en su mayor parte por ácidos grasos con una o varias insaturaciones (poliinsaturados).
Debido a su mecanismo de síntesis, los ácidos grasos naturales tienen un número par de carbonos sin que esto quiera decir que no los haya de números impares, ramificados y sustituidos con grupos funcionales.
Los ácidos grasos más abundantes en la naturaleza son el ácido oleico (~30 % del total de ácidos grasos) y el palmítico que representa por lo general de 10 a 50 % del total de ácidos grasos.

NOMENCLATURA
Nomenclatura sistemática. Los ácidos grasos se denominan de acuerdo al hidrocarburo del que provienen más el sufijo “oico”. El ácido graso de 16 carbonos se llama hexadecanoico.
El nombre común o trivial generalmente relacionado con la fuente natural del cual proviene con la terminación “ico”. El hexadecanoico, se llama más a menudo palmítico porque se obtiene del áceite de palma.
La representación más práctica de los ácidos grasos señala el número de carbonos de la cadena seguido de dos puntos y del número de dobles enlaces por ejemplo: ácido palmítico 16:0 y el ácido oleico 18:1.
En algunos casos se menciona con la letra delta la posición de los dobles enlaces empezando por el carbono del carboxilo, el ácido oleico sería 18:1 delta 9. Si empezamos por el carbono terminal, el mismo ácido sería 18:1 delta 9; el ácido linoleico sería 18:2 delta 9,12 y
18:2 delta 6.
CERAS
Presentes en los vegetales y en los animales marinos, las ceras también se encuentran en los mamíferos como sustancias de protección y en funciones especiales. Las ceras están formadas por un ácido graso de cadena larga, esterificado con un alcohol, también de cadena larga. A diferencia de las grasas no son asimilables por el organismo humano. Las más conocidas son la cera de abeja, la cera de ovejas (la lanolina) y la del aceite de ballena. Son altamente insolubles en agua y son sólidos y duros a temperatura ambiente.

LÍPIDOS COMPUESTOS
Glicerofosfolípidos: Son un grupo numeroso de lípidos compuestos, importantes en la estructura de las membranas y derivados del ácido fosfatídico, que tienen como alcohol al glicerol y que incluyen las lecitinas, cefalinas, plasmalógenos y algunos otros lípidos menos frecuentes.

Lecitinas: También llamadas fosfatidil colinas, están formadas por glicerol, dos ácidos grasos, ácido fosfórico y la base nitrogenada colina. Las lecitinas son los fosfolípidos –así llamados porque contienen fosfato- mas abundantes en las membranas celulares y en el plasma sanguíneo.
Cefalinas: También se llaman fosfatidil serinas y fosfatidil etanolaminas, son fosfolípidos importantes que contienen un glicerol, dos ácidos grasos, un ácido fosfórico y la base nitrogenada serina o etanolamina.
Plasmalógenos: Son fosfolípidos de estructura muy semejante a las lecitinas y las cefalinas pero que tienen el ácido graso de la posición 1 bajo la forma de un aldehído alfa-beta insaturado. Están formados por un glicerol, un aldehído graso, un ácido graso, ácido fosfórico y una base nitrogenada que puede ser colina, etanolamina o serina.

ESFINGOLÍPIDOS
Son un grupo de lípidos compuestos derivados del alcohol aminado esfingosina. La unidad fundamental de los esfingolípidos está formada por una esfingosina unida en enlace amida con un ácido graso de cadena larga para formar la ceramida, a la cual se une algún grupo polar que sirve de cabeza.

Esfingomielinas
Son los esfingolípidos mas abundantes en la cubierta de mielina de las fibras nerviosas y por tener fosfato se los incluye a menudo dentro del grupo de los fosfolípidos. Están formadas por la esfingosina unida en enlace amida con un ácido graso saturado de cadena larga (ceramida) de mas de 20 carbonos.
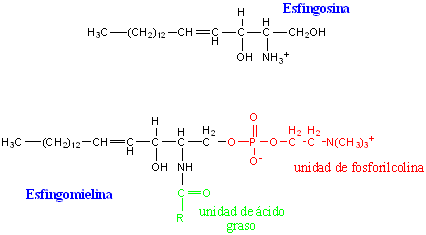
Galactolípidos o cerebrósidos
Al igual que las esfingomielinas, son lípidos complejos, especializados y abundantes en el tejido nervioso. Están formados por una ceramida en enlace glucosídico con el monosacárido galactosa, menos frecuentemente la glucosa o algún oligosacárido (gangliósidos).

LÍPIDOS ASOCIADOS
Los lípidos simples y los compuestos comparten entre sí una de las propiedades más generales de los lípidos, que es la de ser todos ellos ésteres de los ácidos grasos; mientras que los lípidos asociados pueden o no, estar esterificados y se incluyen dentro de la categoría de los lípidos por su naturaleza no polar que los hace solubles en los solventes orgánicos y por salir junto con los lípidos cuando estos se extraen de los tejidos. Según su estructura química, los lípidos asociados pueden dividirse en tres series: terpenoides, eicosanoides y esteroides.
TERPENOIDES
La palabra terpenoide se refiere a una clase muy variada de compuestos similares a los terpenos, una estructura que deriva de la unidad de 5 carbonos llamada isopreno (2-metil-1,3-butadieno) y que tiene un contenido mínimo de 10 átomos de carbono o los más grandes pueden llegar a tener cientos de ellos. Los terpenos de importancia biológica incluyen:
Monoterpenoides: como el limoneno con 2 unidades isoprenoides (10 átomos de carbono). Es el responsable del olor característico de las frutas cítricas.
Sesquiterpenoides: como el farnesol con 3 unidades isoprenoides (15 átomos de carbono). Intermediario en la síntesis de colesterol, precursor de esteriodes.
Diterpenoides: como la vitamina A con 4 unidades isoprenoides (20 átomos de carbono). Asociada al mecanismo de la visión.
Triterpenoides: como el escualeno con 6 unidades isoprenoides (30 átomos de carbono). Intermediario en la síntesis de colesterol, precursor de esteriodes.
Tetraterpenos: como el beta caroteno con 8 unidades isoprenoides (40 átomos de carbono). Fuente del color anaranjado de las zanahorias, precursor de la vitamina A.
Politerpenoides: son los que contienen más de 8 unidades isoprenoides como el dolicol con 19 unidades isoprenoides (95 átomos de carbono). Interviene en la síntesis de peptidoglicanos en células de mamíferos.
El ejemplo más conocido es el de la vitamina A y sus precursores, los carotenos que están presentes en los pigmentos vegetales de color rojo o naranja, como en los jitomates y las zanahorias.
Derivados de terpenos
Incluyen los lípidos derivados de la benzoquinona y del tocol, es decir la coenzima Q, la vitamina K y la vitamina E. Estas moléculas se asocian a menudo con cadenas de unidades isoprenoides más o menos largas, por lo que son los grupos de cabeza los que los distinguen de los Terpenoides. La vitamina K participa como activador en el proceso de la coagulación y una estructura semejante a ella se encuentra en la coenzima-Q que participa en el transporte de electrones y es la molécula más abundante en la cadena respiratoria, se halla provista de una cola hidrofóbica de 10 unidades isoprenoides.

Lípidos
IMPORTANCIA BIOLÓGICA
Los lípidos son biomoléculas orgánicas de distribución prácticamente universal en los seres vivos y que desempeñan en ellos numerosas funciones biológicas, como son:
- Los lípidos constituyen el material fundamental de todas las membranas celulares y subcelulares, en las que aportan la bicapa de fosfolípidos, arreglados con las cabezas polares hacia fuera y las colas no polares hacia dentro.

- Los lípidos forman la mayor reserva de energía de los organismos, que en el caso del organismo humano normal, son suficientes para mantener el gasto energético diario durante la inanición por un período cercano a los 50 días; mientras que el glucógeno corporal alcanza solamente para cerca de 16 horas y las proteínas corporales que teóricamente aportarían casi la misma energía que las grasas, son demasiado importantes para permitir su degradación masiva.

- Las grasas funcionan como aislante térmico muy efectivo para proteger a los organismos del frío ambiental, por lo que los animales de las zonas frías del planeta se protegen con una gruesa capa de grasa bajo la piel y también las grasas sirven de un amortiguador mecánico efectivo, que protege los órganos internos como el corazón y el riñón.

- Los lípidos funcionan como hormonas de gran relevancia para la fisiología humana, por ejemplo las hormonas esteroideas, las prostaglandinas y segundos mensajeros hormonales, como el inositol-trifosfato y también como las vitaminas liposolubles A,D, E y K que forman parte de los lípidos asociados.

- Los lípidos tienen una función nutricional importante y figuran en la dieta tipo aportando alrededor del 30 % de las kilocalorías de la dieta y como fuente de los ácidos grasos indispensables: Linoléico, linolénico y araquidónico.

Enfermedades del metabolismo de carbohidratos
- Intolerancia a la fructosa----> aldolasa hepatica
- Intolerancia a la lactosa----> lactasa
- Galactosemia---> galactosa 1 fosfato uridil transferasa
- Enfermedad de Von Gierke--->glucosa 6 fosfatasa
- Enfermedad de Pompe---> alfa 1-4 glucosidasa
- Enfermedad de Cori---> enzima desramificante del glucogeno
- Enfermedad de Mc Cardle---> fosforilasa muscular
Niveles de glucosa


NIVELES BAJOS DE GLUCOSA (3.3 A 3.9 mmol/L)
•DURANTE EL EMBARAZO Y EN EL NEONATO SE PUEDE PRESENTAR ESTÁ CONCENTRACIÓN DE GLUCOSA EN SANGRE
•EN EL ALCOHOLICO CRÓNICO , SE PRESENTAN ESTOS NIVELES DE GLUCOSA
•CONCENTRACIÓN SANGUÍNEA DE GLUCOSA DURANTE EL AYUNO
•ESTA CONCENTRACIÓN DE GLUCOSA EN SANGRE SE PUEDE PRESENTAR POR UN EXCESO DE INSULINA
•EN ESTA CONDICIÓN SE ESTIMULA LA SECRECIÓN DE GLUCAGÓN
NIVELES NORMALES DE GLUCOSA (4.5 A 5.5 mmol/L)
•LA GLUCOGENÓLISIS Y LA GLUCONEOGÉNESIS CONTRIBUYEN A LOGRAR ESTE ESTADO EN CONDICIONES CUANDO NO SE INGIERE ALIMENTO POR UN LAPSO CORTO (HIPOGLUCEMIA)
NIVELES ALTOS DE GLUCOSA (6.5 A 7.2 mmol/L)
•CONCENTRACIÓN A LA QUE SE ELEVA LA GLUCOSA SANGUÍNEA DESPUÉS DE INGERIR UNA COMIDA RICA EN CARBOHIDRATOS (HIPERGLUCEMIA)
Gluconeogénesis
Cuando los niveles de glucosa son bajos y la reserva de glucógeno esta por agotarse se lleva a cabo esta vía que consiste en la síntesis de glucosa a partir de sustratos no glucocidicos. Esta vía permite que en condiciones de bajos niveles de glucosa los tejidos como cerebro, eritrocitos, cristalino, medula renal, córnea del ojo y testículos










Balance




En los siguientes esquemas se muestran algunos de los alimentadores no glucocidicos




En la siguiente tabla se muestran los aminoácidos que pueden entrar a la gluconeogénesis:

Los aminoácidos lisina y leucina no pueden entrar a la gluconeogénesis
La gluconeogénesis consta de 10 pasos de los cuales siete son iguales a la glucólisis y tres son conocidos como rodeos y son aquellos pasos de la glucólisis en los que se produjo o gasto ATP

Primer rodeo

Segundo rodeo

Tercer rodeo


Regulación alosterica

Regulación hormonal

Suscribirse a:
Entradas (Atom)